Problème n° 10 de l’évolution : le néodarwinisme a une longue histoire de prédictions darwiniennes inexactes sur les organes vestigiaux et l' »ADN poubelle ».
Par Casey Luskin
23 juillet 2020
Bienvenue dans le Top 10 des problèmes scientifiques liés à l’évolution biologique et chimique.
Note de l’éditeur : Il s’agit de la dixième partie d’une série de dix articles basés sur le chapitre de Casey Luskin, « Les dix principaux problèmes scientifiques liés à l’évolution biologique et chimique« , du livre More than Myth, édité par Paul Brown et Robert Stackpole (Chartwell Press, 2014.) Les autres chapitres individuels peuvent être consultés ici : Problème 1, Problème 2, Problème 3, Problème 4, Problème 5, Problème 6, Problème 7, Problème 8, Problème 9, Problème 10, Problème supplémentaire.
Pendant des décennies, les évolutionnistes ont affirmé que nos corps et nos génomes sont remplis de parties et de matériel génétique inutiles – des organes « vestigiaux » – qui montrent que la vie est le résultat d’une évolution non guidée se produisant sur des temps immémoriaux. Lors du procès Scopes en 1925, le biologiste évolutionniste Horatio Hackett Newman affirma qu’il y avait plus de 180 organes et structures vestigiaux dans le corps humain, nombre « suffisant pour faire d’un homme un véritable musée ambulant d’antiquités157. »
Au fil du temps, cependant, ces prédictions concernant les organes vestigiaux du corps et l’ADN inutile ne se sont pas vérifiées. Au fur et à mesure que les scientifiques ont appris à mieux connaître le fonctionnement de la biologie, des fonctions et des finalités importantes ont été découvertes pour ces structures dites vestigiales. En effet, en 2008, la revue New Scientist a rapporté que, depuis l’époque du professeur Newman, la liste des organes vestigiaux « s’est allongée, puis s’est à nouveau réduite » au point qu’aujourd’hui « les biologistes sont extrêmement prudents lorsqu’il s’agit de parler d’organes vestigiaux158. » Les structures qui étaient auparavant – et à tort – considérées comme vestigiales comprennent :
- Les amygdales : à une époque, elles étaient systématiquement enlevées. Aujourd’hui, l’on sait qu’elles ont comme fonction dans le système lymphatique d’aider à combattre les infections159.
- Le coccyx (vertèbre caudale) : de nombreux évolutionnistes prétendent encore que c’est un vestige de la queue de nos supposés ancêtres primates160, mais c’est en fait une partie vitale de notre squelette, utilisée pour attacher les muscles, les tendons et les ligaments qui soutiennent les os de notre bassin.
- La thyroïde : cette glande située dans le cou était autrefois considérée comme sans finalité, et a été ignorée, voire détruite, par les médecins qui opéraient sous de fausses hypothèses darwiniennes. Aujourd’hui, les scientifiques savent qu’elle est vitale pour la régulation du métabolisme.
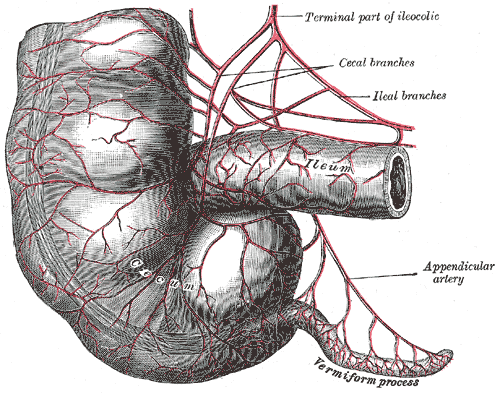
- L’appendice : les scientifiques darwiniens ont affirmé que l’appendice est un « vestige de nos ancêtres herbivores161 » et que, au cours de l’évolution, sa fonction chez l’homme a été diminuée, voire perdue. Mais l’on sait aujourd’hui que l’appendice remplit des fonctions importantes, comme celles de réservoir de bactéries bénéfiques et de production de globules blancs, et joue un rôle important dans le développement du fœtus162. A la lumière de ces preuves, l’immunologiste William Parker de l’université Duke a observé que « de nombreux textes de biologie font encore référence à l’appendice comme à un « organe vestigial », mais « il est temps de corriger les manuels163. »
Malgré le fait attesté déplorable que les biologistes évolutionnistes ont toujours affirmé que les organes n’étaient que des vestiges, ils ont appliqué ce même type de réflexion à nos génomes. Beaucoup ont postulé que la nature aléatoire des mutations remplirait nos génomes de déchets génétiques inutiles, surnommés « ADN poubelle ». Cette hypothèse semble avoir été confirmée lorsque l’on a découvert que seulement 2 % du génome humain interviennent dans le codage des protéines, laissant les 98 % restants inexpliqués. De nombreux scientifiques qui se font les porte-paroles de la biologie évolutive ont affirmé que cette preuve constitue la preuve même de l’évolution darwinienne qui clôt le dossier :
- Kenneth Miller, biologiste évolutionniste de l’université Brown, affirme que « le génome humain est truffé de pseudogènes, de fragments de gènes, de gènes ‘orphelins’, d’ADN ‘poubelle’ et de tant de copies répétées de séquences d’ADN inutiles qu’il ne peut être attribué à rien qui ressemble à un dessein intelligent164. »
- Richard Dawkins écrit également que « les créationnistes pourraient passer un certain temps à spéculer sur les raisons pour lesquelles le Créateur devrait se donner la peine de remplir les génomes de pseudogènes non traduits et d’ADN poubelle répété en tandem165. »
- Dans son livre The Language of God, publié en 2006, Francis Collins affirme qu’environ « 45 % du génome humain » sont constitués de « flotsam et jetsam génétiques166 » (des flotsam et jetsam, bien sûr, sont des déchets inutiles flottant sur l’océan.) Se rapprochant de Dawkins, il en précise les implications :
« À moins que l’on ne soit prêt à adopter la position selon laquelle Dieu a placé [de l’ADN répétitif non fonctionnel] dans ces positions précises pour nous confondre et nous induire en erreur, la conclusion d’un ancêtre commun pour les humains et les souris est pratiquement inévitable167. »
Le problème avec ces arguments n’est pas tant théologique que scientifique : de nombreux exemples de fonction ont été découverts pour ce que l’on appelle l’ADN poubelle.
Le biologiste Richard Sternberg a étudié la littérature et a trouvé de nombreuses preuves d’une fonctionnalité pour l’ADN répétitif. Dans les Annals of New York Academy of Sciences, il a découvert que les fonctions des répétitions comprennent la formation de structures nucléaires d’ordre supérieur, de centromères, de télomères et de centres de nucléation pour la méthylation de l’ADN. Il a été constaté que l’ADN répétitif était impliqué dans la prolifération des cellules, les réponses au stress cellulaire, la traduction des gènes et la réparation de l’ADN168. Sternberg a conclu que « le récit égoïste de l’ADN et les cadres connexes doivent rejoindre les autres ‘icônes’ de la théorie de l’évolution néodarwinienne qui, malgré leurs divergences avec les preuves empiriques, persistent néanmoins dans la littérature169. »
D’autres recherches ont continué à découvrir des fonctions pour divers types d’ADN répétitif, notamment les éléments SINE170, LINE171 et Alu172. Un article a même suggéré que des séquences Alu répétitives pourraient intervenir dans « le développement de fonctions cérébrales supérieures » chez l’homme173. De nombreuses autres fonctions ont été découvertes pour divers types d’ADN non codant pour les protéines, notamment :
- la réparation de l’ADN174,
- l’aide à la réplication de l’ADN175,
- la régulation de la transcription de l’ADN176,
- l’aide au repliement et au maintien des chromosomes177,
- le contrôle de l’édition et de l’épissage de l’ARN178,
- la contribution à la lutte contre les maladies179,
- la régulation du développement embryonnaire180.
Sternberg, ainsi que le généticien James Shapiro de l’université de Chicago, ont prédit en 2005 dans la revue Cytogenetic and Genome Research qu' »un jour, nous considérerons ce que l’on appelait autrefois ‘l’ADN poubelle’ comme un élément essentiel de régimes de contrôle cellulaire véritablement ‘experts’181. »
Le jour prévu par Sternberg et Shapiro est peut-être arrivé plus tôt que prévu. En septembre 2012, la revue Nature a publié les résultats d’un projet de recherche qui a duré un an, rassemblant plus de 400 scientifiques internationaux qui étudient les fonctions de l’ADN non codant chez l’homme. Baptisé le projet ENCODE, cet ensemble de 30 articles novateurs a montré que la « majeure partie » du génome est fonctionnelle. L’article principal faisant état des résultats d’ENCODE affirmait ce qui suit :
« Ces données nous ont permis d’attribuer des fonctions biochimiques à 80 % du génome, en particulier en dehors des régions codant pour les protéines bien étudiées182. »
Ewan Birney, le coordinateur principal de l’analyse des données d’ENCODE a commenté dans le magazine Discover que puisqu’ENCODE n’a examiné que 147 types de cellules, et que le corps humain en compte quelques milliers, « il est probable que 80 % d’entre elles passent à 100 %183. » Le même article a cité Tom Gingeras, un scientifique en chef d’ENCODE, qui a noté que « presque chaque nucléotide est associé à une fonction d’une sorte ou d’une autre, et nous savons maintenant où ils se trouvent, ce qui les lie, quelles sont leurs associations, et plus encore184. » Un autre commentaire dans Nature a noté que « 80 % du génome contiennent des éléments liés à des fonctions biochimiques, ce qui dissipe l’opinion largement répandue selon laquelle le génome humain est essentiellement de l »ADN poubelle’185. » Le Discover Magazine l’a exprimé ainsi : « L’idée essentielle est : ce n’est pas de l »ADN poubelle’186. »
Bien qu’il y ait encore beaucoup de choses que nous ne savons pas sur le génome, la tendance de la recherche pointe clairement dans une direction : plus nous étudions le génome, plus nous détectons des fonctions pour l’ADN non codant. Pourtant, le paradigme de « l’ADN poubelle », désormais controversé, est né et s’est développé dans le cadre du paradigme de l’évolution fondé sur l’idée selon laquelle notre génome a été construit par des mutations aléatoires. Oui, quelques biologistes malhonnêtes ont osé chercher une fonction pour l’ADN non codant, mais la vision darwinienne de la génétique de « l’ADN poubelle » a généralement entravé les progrès de la science, comme l’a admis un article paru en 2003 dans Science :
« Bien qu’accrocheur, l’expression ‘ADN poubelle’ a longtemps repoussé les chercheurs traditionnels dans l’étude de l’ADN non codant. Qui, à part un petit nombre de clochards de la génomique, voudrait fouiller dans les déchets de la génomique ? Cependant, en science comme dans la vie normale, il existe des clochards qui, au risque d’être ridiculisés, explorent des territoires impopulaires. À cause d’eux, la vision de l’ADN poubelle, en particulier des éléments répétitifs, a commencé à changer au début des années 1990. Aujourd’hui, de plus en plus de biologistes considèrent les éléments répétitifs comme un trésor génomique187. »
En dépit des hypothèses darwiniennes répandues selon lesquelles le contraire serait vrai, l’article conclut que « les éléments répétitifs ne sont pas de l’ADN poubelle inutile, mais plutôt des composants importants et intégraux188 » des génomes animaux. Des études suggèrent que ces longues portions d’ADN non codant entre les gènes « constituent une couche importante de régulation du génome dans un large éventail d’espèces189. »
Comme les éléments répétitifs, les pseudogènes sont un autre type d’ADN « poubelle » dont on découvre la fonctionnalité. L’on pense que les pseudogènes sont des copies de gènes autrefois fonctionnels qui ont été inactivés par des mutations. Un article paru dans Science Signaling observe que « les pseudogènes ont longtemps été considérés comme de l’ADN poubelle190« , mais note que les pseudogènes sont des copies de gènes autrefois fonctionnels qui ont été inactivés par des mutations :
« Des avancées récentes ont établi que l’ADN d’un pseudogène, l’ARN transcrit d’un pseudogène ou la protéine traduite d’un pseudogène peuvent avoir des fonctions multiples et diverses et que ces fonctions peuvent affecter non seulement leurs gènes parents, mais aussi des gènes non apparentés. Par conséquent, les pseudogènes sont apparus comme une classe de modulateurs sophistiqués de l’expression des gènes, jusqu’alors peu considérés, avec une implication multiforme dans la pathogenèse du cancer humain191. »
En effet, les fonctions de nombreux pseudogènes ont déjà été découvertes192 ; le projet ENCODE à lui seul a trouvé plus de 850 pseudogènes qui sont « transcrits et associés à la chromatine active193. » Mais que font exactement ces pseudogènes ? Un article paru en 2011 dans la revue RNA affirme à nouveau qu’ils peuvent réguler l’expression des gènes :
« Les pseudogènes ont longtemps été qualifiés d’ADN ‘poubelle’, des copies ratées de gènes qui apparaissent au cours de l’évolution des génomes. Cependant, des résultats récents remettent en question cette appellation ; en effet, certains pseudogènes semblent receler le potentiel de réguler leurs cousins codant pour des protéines194. »
De même, un article publié en 2012 dans la revue RNA Biology déclarait que « les pseudogènes ont longtemps été considérés comme de l’ADN poubelle génomique », mais que « la régulation des pseudogènes est très fréquente195 » dans les organismes multicellulaires complexes. L’article suggérait l’hypothèse selon laquelle « la grande abondance et la conservation des pseudogènes dans une multitude d’espèces indiquent que des pressions sélectives préservent ces éléments génétiques et suggèrent qu’ils peuvent effectivement remplir des fonctions biologiques importantes196. »
Les pseudogènes sont un autre bon exemple de la façon dont les biologistes darwiniens ont supposé qu’un type d’ADN non codant qu’ils ne comprenaient pas était un déchet génétique sans fonction, et ont donc ignoré leurs fonctions. En effet, l’article précité dans RNA Biology explique qu’une des raisons pour lesquelles les évolutionnistes ont été si lents à abandonner l’hypothèse selon laquelle les pseudogènes sont des déchets est que leurs fonctions sont difficiles à détecter. Les auteurs observent que « presque tous les pseudogènes qui présentent une activité biologique significative sont exprimés dans des tissus ou des lignées cellulaires spécifiques », ce qui signifie que seuls des tissus ou des lignées cellulaires spécifiques peuvent utiliser un pseudogène donné pour une certaine fonction. En outre, il est difficile de détecter la fonction des pseudogènes car nous n’avons pas les outils de recherche nécessaires pour comprendre comment ils influencent l’expression des gènes. L’article prédit que « de plus en plus de pseudogènes fonctionnels seront découverts au fur et à mesure du développement de nouvelles technologies biologiques » et conclut que « l’étude des pseudogènes fonctionnels n’en est qu’à ses débuts197. » En effet, deux biologistes de premier plan qui écrivent dans l’Annual Review of Genetics ont indiqué que « les pseudogènes qui ont été convenablement étudiés ont souvent des rôles fonctionnels198. »
De nombreux biologistes évolutionnistes sont convaincus que nos génomes sont pleins de déchets et résistent à l’interprétation selon laquelle pratiquement tout l’ADN est fonctionnel. En effet, un manuel d’évolution de 2012 enseigne que « plus de la moitié du génome n’est composée ni de gènes, ni de vestiges de gènes humains, ni de régions régulatrices. Au contraire, elle est constituée de segments d’ADN semblables à des parasites199… » Pendant ce temps, les preuves continuent de pointer dans la direction opposée. Bien qu’il en reste beaucoup à apprendre sur le fonctionnement de notre génome, la tendance de la recherche est sans ambiguïté : plus nous étudions l’ADN non codant, plus nous trouvons des preuves de fonctionnalité sur de vastes portions de l’ADN.
Références :
157 Horatio Hackett Newman, cité dans The World’s Most Famous Court Trial: Tennessee Evolution Case, 2e éd. (Dayton, TN : Bryan College, 1990), 268. Voir également Robert Wiedersheim, The Structure of Man: An Index to His Past History (Londres : MacMillan and Co, 1895 ; réimprimé par Kessinger, 2007.)
158 Laura Spinney, « Vestigial organs: Remnants of evolution », New Scientist, 2656 (14 mai 2008), à l’adresse http://www.newscientist.com/article/mg19826562.100-vestigial-organs-remnants-of-evolution.html.
159 Sylvia S. Mader, Inquiry into Life, 10e édition (McGraw Hill, 2003), 293.
160 Laura Spinney, « The Five things humans no longer need », New Scientist (19 mai 2008), à l’adresse http://www.newscientist.com/article/dn13927-five-things-humans-no-longer-need.html.
161 Douglas Theobald, « 29+ Evidences for Macroevolution », TalkOrigins.org, à http://www.talkorigins.org/faqs/comdesc/section2.html.
162 Voir Loren G. Martin, « Quelle est la fonction de l’appendice humain ? Scientific American (21 octobre 1999), à l’adresse suivante : http://www.scientificamerican.com/article.cfm?id=what-is-the-function-of-t.
163 William Parker a cité Charles Q. Choi, « The Appendix » : The Appendix : Useful and in Fact Promising », LiveScience (24 août 2009.)
164 Miller, « Life’s Grand Design », 24-32. Miller cite les « gènes orphelins », mais ceux-ci ne sont normalement pas considérés comme des gènes non fonctionnels. Les gènes orphelins sont plutôt des gènes fonctionnels qui n’ont aucune homologie connue avec un autre gène. Ces gènes orphelins constituent la preuve d’une conception intelligente car il n’y a pas de source plausible produisant leurs informations.
165 Richard Dawkins, « The Information Challenge », The Skeptic, 18 (décembre 1998.)
166 Francis Collins, The Language of God: A Scientist Presents Evidence for Belief (New York : Free Press, 2006), 136-37.
167 Ibid, pp. 134-137.
168 Richard Sternberg, « On the Roles of Repetitive DNA Elements in the Context of a Unified Genomic- Epigenetic System », Annals of the New York Academy of Sciences, 981 (2002) : 154-88.
169 Ibid.
170 Ibid.
171 Tammy A. Morrish, Nicolas Gilbert, Jeremy S. Myers, Bethaney J. Vincent, Thomas D. Stamato, Guillermo E. Taccioli, Mark A. Batzer et John V. Mora « DNA repair mediated by endonuclease-independent LINE-1 retrotransposition », Nature Genetics, 31 (juin 2002) : 159-65.
172 Galit Lev-Maor, Rotem Sorek, Noam Shomron et Gil Ast, « The birth of an alternatively spliced exon : 3′ splice-site selection in Alu exons », Science, 300 (23 mai 2003) : 1288-91 ; Wojciech Makalowski, « Not junk after all », Science, 300 (23 mai 2003) : 1246-47.
173 Nurit Paz-Yaacova, Erez Y. Levanonc, Eviatar Nevod, Yaron Kinare, Alon Harmelinf, Jasmine Jacob-Hirscha, Ninette Amariglioa, Eli Eisenbergg et Gideon Rechavi, « Adenosine-to-inosine RNA editing shapes transcriptome diversity in primates », Proceedings of the National Academy of Sciences USA, 107 (6 juillet 2010) : 12174-79.
174 Morrish et coll., « DNA repair mediated by endonuclease-independent LINE-1 retrotransposition », 159-65 ; Annie Tremblay, Maria Jasin et Pierre Chartrand, « A Double-Strand Break in a Chromosomal LINE Element Can Be Repaired by Gene Conversion with Various Endogenous LINE Elements in Mouse Cells », Molecular and Cellular Biology, 20 (janvier 2000) : 54-60 ; Ulf Grawunder, Matthias Wilm, Xiantuo Wu, Peter Kulesza, Thomas E. Wilson, Matthias Mann et Michael R. Lieber, « Activity of DNA ligase IV stimulated by complex formation with XRCC4 protein in mammalian cells », Nature, 388 (31 juillet 1997) : 492-95 ; Thomas E. Wilson, Ulf Grawunder et Michael R. Lieber, « Yeast DNA ligase IV mediates non-homologous DNA end joining », Nature, 388 (31 juillet 1997) : 495-98.
175 Richard Sternberg et James A. Shapiro, « How repeated retroelements format genome function », Cytogenetic and Genome Research, 110 (2005) : 108-116.
176 Jeffrey S. Han, Suzanne T. Szak et Jef D. Boeke, « Transcriptional disruption by the L1 retrotransposon and implications for mammalian transcriptomes », Nature, 429 (20 mai 2004) : 268-74 ; Bethany A. Janowski, Kenneth E. Huffman, Jacob C. Schwartz, Rosalyn Ram, Daniel Hardy, David S. Shames, John D. Minna et David R. Corey, « Inhibiting gene expression at transcription start sites in chromosomal DNAs with antigene RNAs », Nature Chemical Biology, 1 (septembre 2005) : 216-22 ; J. A. Goodrich et J. F. Kugel, « Non-coding-RNA regulators of RNA polymerase II transcription », Nature Reviews Molecular and Cell Biology, 7 (août 2006) : 612-16 ; L.C. Li, S. T. Okino, H. Zhao, H., D. Pookot, R. F. Place, S. Urakami, H. Enokida et R. Dahiya, « Small dsRNAs induce transcriptional activation in human cells », Proceedings of the National Academy of Sciences USA, 103 (14 novembre 2006) : 17337-42 ; A. van de Lagemaat, J. R. Landry et D. L. Mager, P. Medstrand, « Transposable elements in mammals promote regulatory variation and diversification of genes with specialized functions », Trends in Genetics, 19 (octobre 2003) : 530-36 ; S. R. Donnelly, T. E. Hawkins et S. E. Moss, « A Conserved nuclear element with a role in mammalian gene regulation », Human Molecular Genetics, 8 (1999) : 1723-28 ; C. A. Dunn, P. Medstrand et D. L. Mager, « An endogenous retroviral long terminal repeat is the dominant promoter for human B1,3- galactosyltransferase 5 in the colon », Proceedings of the National Academy of Sciences USA, 100 (28 octobre 2003) : 12841-46 ; B. Burgess-Beusse, C. Farrell, M. Gaszner, M. Litt, V. Mutskov, F. Recillas-Targa, M. Simpson, A. West et G. Felsenfeld, « The insulation of genes from external enhancers and silencing chromatin », Proceedings of the National Academy of Sciences USA, 99 (10 décembre 2002) : 16433-37 ; P. Medstrand, Josette-Renée Landry et D. L. Mager, « Long Terminal Repeats Are Used as Alternative Promoters for the Endothelin B Receptor and Apolipoprotein C-I Genes in Humans », Journal of Biological Chemistry, 276 (19 janvier 2001) : 1896-1903 ; L. Mariño-Ramíreza, K. C. Lewisb, D. Landsmana et I. K. Jordan, « Transposable elements donate lineage-specific regulatory sequences to host genomes », Cytogenetic and Genome Research, 110 (2005):333-41.
177 S. Henikoff, K. Ahmad, H. et S. Malik, « The Centromere Paradox: Stable Inheritance with Rapidly Evolving DNA », Science, 293 (10 août 2001) : 1098-1102 ; C. Bell, A. G. West et G. Felsenfeld, « Insulators and Boundaries: Versatile Regulatory Elements in the Eukaryotic Genome », Science, 291 (19 janvier 2001) : 447-50 ; M.-L. Pardue et P. G. DeBaryshe, « Drosophila telomeres: two transposable elements with important roles in chromosomes », Genetica, 107 (1999) : 189-96 ; S. Henikoff, « Heterochromatin function in complex genomes », Biochimica et Biophysica Acta, 1470 (février 2000) : O1-O8 ; L. M. Figueiredo, L. H. Freitas-Junior, E. Bottius, Jean-Christophe Olivo-Marin et A. Scherf, « A central role for Plasmodium falciparum subtelomeric regions in spatial positioning and telomere length regulation », The EMBO Journal, 21 (2002) : 815-24 ; Mary G. Schueler, Anne W. Higgins, M. Katharine Rudd, Karen Gustashaw et Huntington F. Willard, « Genomic and Genetic Definition of a Functional Human Centromere », Science, 294 (5 octobre 2001) : 109-15.
178 Ling-Ling Chen, Joshua N. DeCerbo, et Gordon G. Carmichael, « Alu element-mediated gene silencing », The EMBO Journal 27 (2008) : 1694-1705 ; Jerzy Jurka, « Evolutionary impact of human Alu repetitive elements », Current Opinion in Genetics & Development, 14 (2004) : 603-8 ; G. Lev-Maor et coll., « The birth of an alternatively spliced exon: 3′ splice-site selection in Alu exons », 1288-91 ; E. Kondo-Iida, K. Kobayashi, M. Watanabe, J. Sasaki, T. Kumagai, H. Koide, K. Saito, M. Osawa, Y. Nakamura et T. Toda, « Novel mutations and genotype-phenotype relationships in 107 families with Fukuyama-type congenital muscular dystrophy (FCMD) », Human Molecular Genetics, 8 (1999) : 2303-09 ; John S. Mattick et Igor V. Makunin, « Non-coding RNA », Human Molecular Genetics, 15 (2006) : R17-R29.
179 M. Mura, P. Murcia, M. Caporale, T. E. Spencer, K. Nagashima, A. Rein et M. Palmarini, « Late viral interference induced by transdominant Gag of an endogenous retrovirus », Proceedings of the National Academy of Sciences USA, 101 (27 juillet 2004) : 11117-22 ; M. M. Mura, P. Murcia, M. Caporale, T. E. Spencer, K. Nagashima, A. Rein et M. Palmarini, « Late viral interference induced by transdominant Gag of an endogenous retrovirus », Proceedings of the National Academy of Sciences USA, 101 (27 juillet 2004) : 11117-22 ; M. Kandouz, A. Bier, G. D Carystinos, M. A Alaoui-Jamali et G. Batist, « Connexin43 pseudogene is expressed in tumor cells and inhibits growth », Oncogene, 23 (2004):4763-70.
180 K. A. Dunlap, M. Palmarini, M. Varela, R. C. Burghardt, K. Hayashi, J. L. Farmer et T. E. Spencer, « Endogenous retroviruses regulate periimplantation placental growth and differentiation », Proceedings of the National Academy of Sciences USA, 103 (26 septembre 2006):14390-95 ; L. Hyslop, M. Stojkovic, L. Armstrong, T. Walter, P. Stojkovic, S. Przyborski, M. Herbert, A. Murdoch, T. Strachan et M. Lakoa, « Downregulation of NANOG Induces Differentiation of Human Embryonic Stem Cells to Extraembryonic Lineages », Stem Cells, 23 (2005) : 1035-43 ; E. Peaston, A. V. Evsikov, J. H. Graber, W. N. de Vries, A. E. Holbrook, D. Solter et B. B. Knowles, « Retrotransposons Regulate Host Genes in Mouse Oocytes and Preimplantation Embryos », Developmental Cell, 7 (octobre 2004) : 597-606.
181 Richard Sternberg et James A. Shapiro, « How repeated retroelements format genome function », Cytogenetic and Genome Research, 110 (2005) : 108-116.
182 Le consortium du projet ENCODE, « An integrated encyclopedia of DNA elements in the human genome« , Nature, 489:57-74 (6 septembre 2012.)
183 Ewan Birney, cité dans Ed Yong, « ENCODE: the rough guide to the human genome », Discover Magazine (5 septembre 2012), à l’adresse http://blogs.discovermagazine.com/notrocketscience/2012/09/05/encode-the-rough-guide-to-the-human-genome/.
184 Tom Gingeras, cité dans Ed Yong, « ENCODE: the rough guide to the human genome », Discover Magazine (5 septembre 2012), à l’adresse http://blogs.discovermagazine.com/notrocketscience/2012/09/05/encode-the-rough-guide-to-the-human-genome/.
185 Joseph R. Ecker, « Serving up a genome feast », Nature, 489:52-55 (6 septembre 2012.)
186 Ed Yong, « ENCODE: the rough guide to the human genome », Discover Magazine (5 septembre 2012), à l’adresse http://blogs.discovermagazine.com/notrocketscience/2012/09/05/encode-the-rough-guide-to-the-human-genome/.
187 Makalowski, « Not Junk After All », 1246-1247.
188 Ibid.
189 David R. Kelley et John L. Rinn, « Transposable elements reveal a stem cell specific class of long noncoding RNAs », Genome Biology, 13:R107 (2012.)
190 Laura Poliseno, « Pseudogenes: Pseudogenes: Newly Discovered Players in Human Cancer », Science Signaling, 5 (242) (18 septembre 2012.)
191 Ibid.
192 Voir par exemple D. Zheng et M. B. Gerstein, « The ambiguous boundary between genes and pseudogenes: the dead rise up, or do they ? », Trends in Genetics, 23 (mai 2007) : 219-24 ; S. Hirotsune et coll., « An expressed pseudogene regulates the messenger-RNA stability of its homologous coding gene », Nature, 423 (1er mai 2003) : 91-96 ; O. H. Tam et al, « Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes », Nature, 453 (22 mai 2008) : 534-38 ; D. Pain et coll., « Multiple Retropseudogenes from Pluripotent Cell-specific Gene Expression Indicates a Potential Signature for Novel Gene Identification », The Journal of Biological Chemistry, 280 (25 février 2005) : 6265-68 ; J. Zhang et coll., « NANOGP8 is a retrogene expressed in cancers », FEBS Journal, 273 (2006) : 1723-30.
193 Le consortium du projet ENCODE, « An integrated encyclopedia of DNA elements in the human genome« , Nature, 489:57-74 (6 septembre 2012.)
194 Ryan Charles Pink, Kate Wicks, Daniel Paul Caley, Emma Kathleen Punch, Laura Jacobs et David Paul Francisco Carter, « Pseudogenes : Pseudo-functional or key regulators in health and disease ? », RNA, 17 (2011) : 792-798.
195 Yan-Zi Wen, Ling-Ling Zheng, Liang-Hu Qu, Francisco J. Ayala et Zhao-Rong Lun, « Pseudogenes are not pseudo no more », RNA Biology, 9(1):27-32 (janvier 2012.)
196 Ibid.
197 Ibid.
198 Evgeniy S. Balakirev et Francisco J. Ayala, « Pseudogenes, Are They ‘Junk’ or Functional DNA ? », Annual Review of Genetics, 37 (2003) : 123-151.
199 Carl Zimmer et Douglas Emlen, Evolution: Making Sense of Life, p. 132 (Roberts and Company, 2012.)
200 Nicholas Wade, « An Evolutionary Theory of Right and Wrong », The New York Times (31 octobre 2006), consulté le 28 avril 2012, http://www.nytimes.com/2006/10/31/health/psychology/31book.html.
201 Jeffrey P. Schloss, « Evolutionary Accounts of Altruism & the Problem of Goodness by Design » dans Mere Creation ; Science, Faith & Intelligent Design, édité par William A. Dembski (Downers Grove, IL, Intervarsity Press, 1998), 251.
202 Francis Collins cité dans Dan Cray, « God vs. Science », Time Magazine (5 novembre 2006), consulté le 28 avril 2012, http://www.time.com/time/printout/0,8816,1555132,00.html.
203 Ibid.
204 Jeffrey P. Schloss, « Emerging Accounts of Altruism: ‘Love Creation’s Final Law' », dans Altruism and Altruistic Love: Science, Philosophy, & Religion dans Dialogue, éditeurs. Stephen G. Post, Lynn G. Underwood, Jeffrey P. Schloss et William B. Hurlbut (Oxford : Oxford University Press, 2002), 221.
205 Philip S. Skell, « Why do we invoke Darwin ? », The Scientist, 19 (29 août 2005) : 10.
206 Noam Chomsky, Language and Mind, 3e édition (Cambridge : Cambridge University Press, 2006), 59.
L’article original en anglais a été publié sur Evolution News à l’adresse https://www.discovery.org/a/24041/#fn142 le 20 février 2015 et a été traduit en français et republié sur Bible & Science Diffusion avec autorisation.
A propos de Casey Luskin

Casey Luskin
Casey Luskin est scientifique et avocat, et titulaire de diplômes d’études supérieures en sciences et en droit. Il a obtenu une licence et une maîtrise en sciences de la terre à l’université de Californie à San Diego, où il a beaucoup étudié la géologie et l’évolution, tant au niveau du premier que du deuxième cycles. Sa thèse de maîtrise portait sur le paléomagnétisme de la plaine de la rivière Snake dans le Sud de l’Idaho.
Depuis 2005, il est avocat agréé en Californie, après avoir obtenu un diplôme de droit à la faculté de droit de l’université de San Diego, où ses études ont porté sur le droit du Premier amendement de la Constitution, le droit de l’éducation et le droit de l’environnement. Il a également mené des recherches géologiques à la Scripps Institution for Oceanography.
En 2001, il a cofondé le centre IDEA (Intelligent Design and Evolution Awareness), une organisation à but non lucratif qui aide les étudiants dans leurs recherches sur le dessein intelligent (ID) en créant des « clubs IDEA » sur les campus des universités et des lycées du monde entier.
De 2005 à 2015, il a travaillé pour le Centre pour la science et la culture du Discovery Institute, d’abord comme responsable de programme en politique publique et affaires juridiques (2005-2010), puis comme coordinateur de recherche (2011-2015.) Dans ces fonctions, il a aidé et défendu des scientifiques, des éducateurs et des étudiants qui cherchaient à étudier, effectuer des recherches et enseigner librement sur le débat scientifique concernant l’évolution et l’identité néodarwiniennes. Comme expliqué sur son site personnel, au 31 décembre 2015, il ne travaille plus comme membre du personnel du Discovery Institute car il a pour objectif de poursuivre ses études.
Certaines de ses publications sont parues dans des revues techniques de droit et de sciences et dans d’autres revues spécialisées, notamment le Journal of Church and State ; la Montana Law Review ; la Hamline Law Review ; la Liberty University Law Review ; la University of St. Thomas Journal of Law & Public Policy ; et Geochemistry, Geophysics, and Geosystems (G3.) Il a également coécrit ou contribué à de nombreux ouvrages.
Il s’intéresse tout particulièrement à la géologie, à l’enseignement des sciences, aux origines biologiques et à la protection de l’environnement.
Formation :
- Doctorat en droit, Université de San Diego.
- Maîtrise en sciences de la terre, Université de Californie, San Diego.
- Licence en sciences de la terre, université de Californie, San Diego.
Affiliations professionnelles passées et/ou présentes :
- Barreau de Californie.
- Association américaine pour l’avancement de la science.
- Association du barreau américain.
- Union géophysique américaine.
- Société scientifique chrétienne.
- Société juridique chrétienne.
- American Scientific Affiliation.